|
 |
 |
 |
 |
 |
 |
 |
|
|
Natural History Articles
Bull.
Mus. Comp. Zool., Harvard Univ., 132 (3); 245-273, December, 1964 No. 3-The Lesser Antillean
Representatives of Bothrops and Constrictor By JAMES D. LAZELL, JR. The
taxonomy of four Lesser Antillean populations of snakes, two of Bothrops
and two of Constrictor, has long been in a state of confusion. This
situation is perhaps explicable, as so many similar situations in West
Indian herpetology are, because there were until now very few examples of
these populations in collections, and there was virtually no record of the
animals in life. I have
combined in this paper a revision of the Lesser Antillean populations of
the two genera because their zoogeography is quite comparable, while their
taxonomy presents two very different patterns of differentiation in island
forms. CONSTRICTOR The
nomenclatural problem in this genus is not, perhaps, less muddled than it
was in Bothrops, but it is certainly easier to untangle.
There is only one South American species, and its nominate
subspecies, Constrictor constrictor
constrictor, is the one closest geographically to the Lesser Antillean
forms, occurring both in Trinidad and Tobago.
As opposed to Bothrops atrox, which I cannot effectively define in relation to the
other species in the genus (except to define the others, and then merely
say that anything not fitting those definitions is a "Bothrops atrox"), c. c. constrictor may be defined for
comparative purposes as follows: Constrictor
constrictor constrictor has
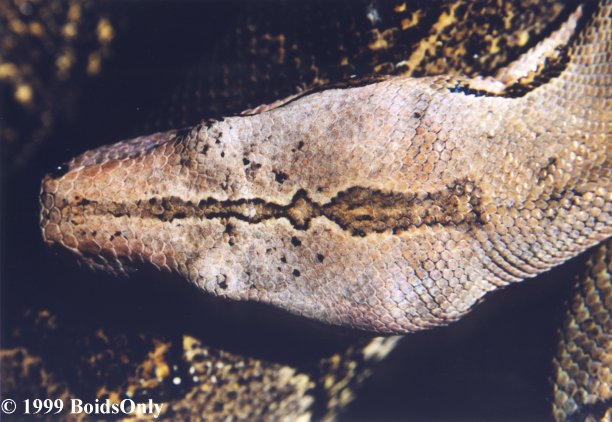
a slightly prominent snout and a straight or nearly straight canthus;
there are 81 to 95 dorsal scale rows at midbody, 234 to 243 ventrals, and
15 to 22 rather neat, geometric, dark dorsal saddles to the level of the
anus; the dorsal ground color is fawn-brown to tan; the venter white to
yellow with little dark marking a dark stripe proceeds from the eye
directly to the supralabials (subocular stripe), and a second proceeds
from the eye anteriorly in the loreal region, then curves down onto the
supralabials (loreal stripe) ; both stripes correspond to two dark
blotches on the infralabials and chin. (See Figures 4 and 5.) The
oldest available date for a Lesser Antillean form is Boa
orophias Linneaus (1758) ; the
ventral count given unequivocally assigns the name to the Saint Lucian
form since no other member of the genus has 28I ventrals.
Dumeril and Bibron (1844) applied the name diviniloqua to a
Saint Lucian specimen; this name was originally proposed by Laurenti
(1768). Therefore, diviniloqua, sensu Dumeril and Bibron, is a junior synonym of orophias;
diviniloquus, sensu Laurenti, does not seem to apply to any
Lesser Antillean form, though, of the two (the Saint Lucian or the
Dominican), it could only have applied to orophias.
No name has ever been proposed for the Dominican form, and the
few available specimens have always been considered under orophias. Stull (1935)
combined both Saint Lucian and Dominican forms under orophias and separated this composite from constrictor at the species level. The
analogous zoogeographical situation is clear: in Bothrops
and Constrictor there are
continental forms which occur also on
Trinidad (and, in Constrictor, on Tobago too) there is a population far to the north
on Saint Lucia in each case, and a third population even farther north, in
the case of Constrictor on
Dominica. The taxonomic
situation is sharply different, however: the geographically intermediate Constrictor is, with respect to most characters of diagnostic value,
morphologically intermediate as well.
There is feasible evidence, then, in Constrictor,
for continuity of evolutionary role which simply does not exist in
Bothrops. I therefore
regard the populations of Constrictor
as members of a stepped-cline series demonstrating geographic
variation in a single species and rank these forms as subspecifically
related to each other. The
first population in geographic sequence is the Tet'chien of Saint Lucia: Boa diviniloqua,
Dumeril et Bibron, 1844, Erp. Gen., 6: 515. Boa
diviniloquax, Jan, 1864, Icon. Gen., p. 81, pl. iii. Boa diviniloqua
(part), Boulenger, 1893, Cat. Snakes, Brit. Mus., 1: 118. Constrictor orophias (part), Stull, 1935, Proc. Boston Soc. Nat. Hist., 40
(8): 405. Barbour, 1914, Mus.
Comp. Zool., 44 (2): 329. Type:
A specimen currently in the Museum de Geer with 280
ventrals is taken to be Linnaeus' type, fide Andersson (1899). Type
locality: None
designated; here restricted to Praslin, Saint Lucia. Diagnosis.
A Constrictor with a prominent
snout and a convex canthus; 65 to 75 dorsal scale rows at midbody, 270 to
288 ventrals, and 27 to 31 distinct, subrectangular, dark dorsal saddles
to the level of the anus; dorsal ground color rich brown; venter white
with black or grey spotting pronounced; subocular stripe distinct and
complete; loreal stripe largely obsolete; dark pigment on chin and
infralabials not closely corresponding to the facial stripes. Description.
In general habitus this form closely corresponds to C.
c. constrictor of Trinidad, Tobago, and part of South America.
The principal differences, aside from squamation, are in the darker
dorsal coloration, increase of grey patches on the venter, and the general
decomposition of the bold, regular pattern so characteristic of C.
c. constrictor. Figures 4 and 5 show the facial markings, more prominent
snout, and dorsal pattern of this form compared with those of C.
c. constrictor. As
compared with C. c. constrictor,
orophias shows an increase in the number of dark dorsal saddles, and
their greater irregularity contributes to the anastomosis of some of the
posterior ones. There is a
decrease in number of dorsal scale rows at midbody; however, there is also
an increase in number of ventrals and, with respect to this character, orophias
is not intermediate between constrictor
and nebulosus
subsp. nov. of Dominica. The
character, however, is far from being absolute; merely a higher average
number of ventrals is not, I think, sufficient to claim that this form
represents a distinct species. Nevertheless,
orophias of Saint Lucia, not nebulosus subsp. nov. of
Dominica, has the highest ventral counts in the genus. Habits
and habitat. Large adults are readily collected simply because of
their stationary habits; a specimen seen a week, two weeks, or even a
month previously is usually in the same vicinity and easily relocated when
one goes to collect it. Some,
e.g. MCZ 75848, from Praslin, were said to have been using the same den
sites for many years. Young specimens are very much more "as you find
them"; I collected several while hunting for other things, and most
often found young ones in trees -one as high as twelve meters from the
ground (MCZ 75845). The
Tet'chien is, at any age, rather lethargic; they hiss a great deal when
aroused and will strike savagely, though their accuracy leaves a good deal
to be desired. They never seem to be found in the same places as Bothrops:
for example, though I got both at Fond Citron of Grande Anse, the Constrictor
was in uncultivated woods, the Bothrops
in a coconut grove. Most
estate owners and managers protect the Tet'chien, sometimes even fining
workers for killing one; this practice is based on the belief that the
Tet'chien destroys rats - which it certainly does - though probably not
with anything like the efficiency of the Serpent.
Barbour (1937) claimed that the Tet'chien is "rare" on
Saint Lucia; I could find no evidence of this.
I never made any special attempt to find one, but rather collected
them only when it was easy and convenient.
The people of Saint Lucia certainly do not regard Constrictor
as the least bit rare, though admit that they are not found in the
fantastic concentrations that Bothrops
often are. To anyone
who has ever seen both a mongoose and a Constrictor
- even a young one - the idea that mongooses could exterminate the
latter must seem a shade ludicrous. Range.
The Tet'chien of Saint Lucia occupies the same region as does Bothrops
caribbaeus, almost exactly. It
does occur to higher elevations, however: ca. 350 meters. Size.
The largest specimen I collected, a male from Anse-La-Raye, was
2.365 meters (MCZ 75847); a female from Praslin (MCZ 75848) was 2.305
meters. Both of these are sizable snakes, but not by any means of
maximum size: a considerably larger individual remains contentedly at
Anse-LA-Raye simply because I had no intention of either attempting to
carry it out of the bush or to preserve it even if I got it out. Relationships. The differences between this form and C. c.
constrictor are absolute
with respect to a number of characters (e.g., ventrals, dorsal saddles and
loreal stripe). Were it not
for the fact that the Dominican form is likewise absolutely distinct, and
even more extreme in the same
direction with respect to all characters except ventral count - so
rendering orophias intermediate
between two extremes of coloration, morphology, and geography - I would
regard full species status as necessitated for this form.
As it happens, however, we have a remarkable stepped-cline
series of which orophias is
the middle member of three steps. Actually,
in terms of color and shape of dorsal markings, orophias
and constrictor are
quite similar; with respect to squamation and number of dorsal markings, orophias
is more similar to the Dominican Tet'chien.
In allusion to the extremely dark, clouded appearance of the latter
form I describe it as: Boa
diviniloqua (part), Boulenger, 1893, Cat. Snakes, Brit. Mus., 1: 118. Constrictor orophias (part), Barbouir, 1914, Mem.
Mus. Comp. Zool., 44 (2): 329; Stull, 1935, Proc. Boston Soc. Nat.
Hist., 40 (8): 405. Type:
MCZ 65493, J. D. Lazell, Jr. coll., 30 July, 1959. Type
locality: Woodford
Hill., Dominica. Diagnosis.
A Constrictor with a prominent
snout and a strikingly convex canthus; 59 to 69 dorsal scale rows at
midbody, 258 to 273 ventrals, and 32 to 35 very obscure, irregular
transverse markings to the level of the anus; dorsal ground color very
dark, clouded, grey-brown; venter ash to slate grey, blotched and mottled
with black; both loreal and subocular stripes absent, or, at most,
partially indicated and largely obsolete infralabials and chin merely grey. Description
of the type. MCZ 65493 is an adult female with 64 dorsal scale
rows at midbody, and 264 ventrals. The
anal is single. There are 19
supralabials on each side; there are 21 infralabials on the right, 20 on
the left. Preserved, the type
measures 1,438 mm, of which 172 mm are tail. There are
33 dark transverse markings to the level of the anus; these markings are
irregular in shape, not much darker than the dorsal ground color and
appear almost more as mottling than actual transverse saddles.
The dorsal ground color, in life, was greyish chocolate-brown.
Posteriorly, the transverse markings become more distinct, black
bordered and dark brown; the ground color, contrastingly, becomes yellower
brown. On the tail,
therefore, the pattern consists of very dark brown, blackbordered saddles
set off by intervals of ochre. The venter
is ash grey anteriorly and becomes very dark -slate grey to black -
posteriorly; there are irregular dark blotches along the lateral edges of
the ventrals anteriorly, and these become obliterated posteriorly.
The chin is entirely grey, though paler medially. The head
has a dark temporal stripe, extending from the eye to beyond the
commissure of the mouth, distinctly darker- bordered along its ventral
edge. In life, there was a
discernible dark streak down the middle of the head. The face is paler
grey, and there was, in life, a pink suffusion below the eye and on the
loreal region. There are no
indications of subocular or loreal stripes. Paratypes:
MCZ 65494, same data as the type; MCZ 65492, Moore
Park, Dominica, J. D. Lazell, Jr. coll., 21 June, 1958; MCZ 65495, Layou
Park, Dominica, J. D. Lazell, Jr. coll., 14 August 1959; MCZ 58772,
Trafalgar, Dominica, J. D. Lazell, Jr. coll., 18 June, 1958; MCZ 74371,
Trafalgar, Dominica, J. D. Lazell, Jr. coll., 18 June, 1959; MCZ 6106,
Portsmouth, Dominica, S. Garman coll., 1879. Variation.
Variation in dorsal scale rows at midbody, number of ventrals, and
number of transverse markings to the level of the anus is tabulated (Table
4). Supralabials vary from 19
to 21; infralabials vary from 20 to 22. The dorsal
ground color varies somewhat, MCZ 65493 representing the light extreme
(still cloudy grey-brown and with very indistinct markings), and MCZ 65492
representing the darkest individual collected (were MCZ 65492 any darker,
I would be forced to call it black).
MCZ 6106, collected in 1879, is pale, but this is due to fading; it
is, despite this, typical in markings and shows them rather well. (A
portion of a head also collected by Garman on the same trip bears the
number MCZ 6107; it is deliberately not designated as a paratype.) Habits
and habitat. Barbour's (1937) comment that this form is "less
uncommon" than the Tet'chien of Saint Lucia is misleading: I would
describe them as being amazingly abundant.
The Dominican Tet'chien, like its relative in Saint Lucia, hisses
loudly when aroused and strikes
rather blindly. Too, it is
basically lethargic and prone to remaining in the same place for long
periods of time. While
staying at Woodford Hill I capitalized on their habitual laziness to save
snake sacks, and merely left the snakes where I found them: they were
almost always still there when I returned, even after several weeks.
Congregations of three to twelve specimens denning in the same
hollow log or tree stump are not at all uncommon, especially along the
edges of machine-cleared banana fields where suitable den sites are often
in profusion. The people
of Dominica, unlike most in Saint Lucia, have no Bothrops
to fear and thus enjoy being deathly afraid of the Tet'chien;
sometimes they even kill them, though usually
only when the snakes take to raiding hen houses, which seems to be
rare. Children, once
initiated, are not at all afraid of even the biggest ones, however, and
their parents are often shamed into changing their minds about the danger
of the Tet'chien simply because the children come to regard them as
play-toys. Accidents
are rare even so: even when a Tet'chien strikes with potentially dangerous
accuracy, it will seldom remember to close its mouth in time to actually
bite. The
Dominican Constrictor, like its
relatives, is often encountered in trees (though large adults rarely are).
Apparently they eat rats almost exclusively now, though agoutis (Dasyproctis)
are abundant in many parts of the island and are no doubt taken as well. Range.
Constrictor constrictor nebulosus seems to occur throughout the island, at least to elevations of ca. 350
meters. It is, however,
confined to wet ravines in dry country, e.g. at the south tip around
Scott's Head, and may be largely replaced along the very dry northern
leeward coast by the "Tet'chien Blanc," Clelia. Size.
The largest specimen collected, MCZ 65490-, is a female of 1,847 mm
total length; the tail is truncated, being only 87 mm long.
Another female, MCZ 65492, is of more normal proportions, being
1,786 mm total length, of which 203 mm is tail.
As with the Saint Lucian form, however, these specimens are far
short of maximum for their taxon. A
specimen owned by Rene Honegger of the Zurich Zoo (one of a litter
produced in 1958 at the Philadelphia Zoo), is now "three meters"
long (pers. comm.). For the sake of practicality, the biggest ones are
best left in the bush. Relationships.
Constrictor constrictor nebulosus is
the terminal form of a stepped-cline series, as discussed under C.
c. orophias. Breeding
experiments with other subspecies of Constrictor
constrictor would be most interesting in assessing whether or not the
subspecific rank I have accorded this form, due to its membership in a
stepped-cline series, has merit from the standpoint of reproductive
potential, as well as geographical and morphological characteristics.
Such experiments are being contemplated by Honegger (pers. comm.),
though the male he intends to use is a specimen of the very different C.
c. occidentalis. It must
be noted, however, that these two completely allopatric forms have never
been subjected to any selection pressure whatever relative to
interbreeding, and therefore the presence or absence of reproductive
isolation between them may be entirely happenstance, if they are
artificially placed in "sympatry." Constrictor constrictor orophias, however, is more different from any other
subspecies of Constrictor
constrictor than any of them are from each other (nebulosus excepted)
with respect to such a conventionally respectable character as ventral
count; this I have nevertheless made a subspecies.
My reasons for ranking these forms as I have center around the
concept of the species set forth by Simpson (1961), in which the principal
criterion of conspecificity is continuity of evolutionary role. A
subspecies, as I would define it, is a rather peculiar sort of
geographical variant within a species that has a wider range; it must be
sharply defined (as opposed to gradually clinal) and diagnostically
homogeneous (as opposed to the sort of situation which arises from
discordant variation). Nevertheless,
subspecies are, from the standpoint of evolutionary role, perfectly
continuous with each other within their species.
Continuity of evolutionary role, even between forms that are
morphologically very dissimilar in many respects, can be maintained by
direct intergradation a d continuous gene flow.
As Simpson (p. -153) points out, however, geographic isolation
tends to break the continuity of evolutionary role by actually breaking
the continuity of gene flow. Therefore,
I am willing to admit as geographic races of a single species only those geographically isolated forms in which there is apparent
continuity of evolutionary role expressed in the characteristics of the
animals. (In the cases
presented, the critical characteristics have been morphological but this
is certainly not obligatory.) In
Constrictor there is a sequential increase proceeding northward
through the three populations in the following, characters: scale size (=
decrease in dorsal scale rows at midbody), number of transverse saddles,
darkness of coloration, irregularity of pattern, obliteration of facial
markings, and convexity of the canthus.
On the other side of the slate is ventral count; in this case the
sequence is broken and the geographically intermediate form has the
highest average. Nevertheless,
the balance of characters examined would seem to indicate that there is a
continuity of evolutionary role, expressed in the morphology of the
populations, proceeding from Constrictor
constrictor constrictor through
Constrictor constrictor orophias
to Constrictor constrictor
nebulosus. In the
case of the Bothrops populations, the geographic conditions are admirably
suited for this sort of pattern, but the animals fail completely to
conform in all discernible characters. Their morphological and behavioral characteristics are
such that by any standard there can be no doubt that the Antillean forms
are distinct species relative to each other.
Assigning one or the other to subspecific rank under one of the
South American forms would require an initial arbitrary choice as to which
of the Antillean populations should be assigned to a South American form -
a subsequent arbitrary choice would then be necessary to determine under
which South American form to place the Antillean population chosen.
In the end, we would be faced with the overwhelming reality that we
had merged forms merely because
they were allopatric. Two
primary objections to my criteria of continuity may be cited: first, the
islands may have been populated in a way which obscures the true
relationships of the animals. That
is, had a distant island been the first place a new form differentiated,
then a second, intermediate one colonized from that stock, and
subsequently differentiated again, in each case the differentiation might
have produced what would have been a stepped-cline series, had the
geographic sequence been intelligible.
Secondly, it is readily seen that no less than three populations
are necessary before we can discern the stepped cline.
That is, if Dominica and its Constrictor
did not exist, I would have classified
orophias as a full species, since there would have been no evidence
for continuity of evolutionary role. Both of
these arguments are quite valid. It must be pointed out, however, that
whenever a geographically and reproductively isolated, absolutely distinct
population is classed as a subspecies a special sort of geographical
variant within a more wide-spread species), this is an act of presumption.
Conservatively speaking, the form qualifies as a species completely. It is
only by means of strong evidence to the contrary that we may really refuse
to rank it as such. As any
taxonomist ultimately [Back To Top] [Back To The Boas] [Main Page]
|